



У раніше не лікованих пацієнтів з діабетом 2 типу метформін змінює кишковий мікробіом, що сприяє терапевтичним ефектам препарату
Завдяки безпеці, низькій вартості, сприятливим ефектам на рівень глюкози крові і кардіоваскулярної смертності метформін є препаратом, що найбільш часто застосовується у фармакотерапії пацієнтів з цукровим діабетом (ЦД) 2 типу. Проте усі механізми його дії залишаються не вивченими.
Вважається, що метформін чинить антигіперглікемічні ефекти, пригнічуючи утворення глюкози в печінці за рахунок активації AMPKзалежних і AMPKнезалежних шляхів (AMPK – АМФактивована протеїнкіназа); проте все більше доказів свідчить, що препарат може діяти в гастроінтестинальному тракті. Наприклад, при пероральному призначенні його глюкозознижуючий ефект більш виражений порівняно з таким при внутрішньовенному використанні. У дослідженнях, в яких порівнювались лікарські форми метформіну зі зниженою і нормальною плазмовою експозицією, було встановлено, що основним місцем дії препарату є нижні відділи кишечнику. Нещодавні дослідження у гризунів і людини також показали, що протидіабетичний ефект метформіну може опосередковуватись змінами мікробіому кишечнику. Втім на сьогодні невідомо, яким чином метформін впливає на кишкову мікрофлору в раніше не лікованих пацієнтів з ЦД 2 типу і як він взаємодіє з кишковими бактеріями.
Метою цього рандомізованого плацебоконтрольованого подвійного сліпого дослідження за участю пацієнтів з уперше діагностованим ЦД 2 типу, що дотримувались обмеженої за калоріями дієти, було вивчити вплив метформіну на склад і функції кишкової мікрофлори за допомогою метагеноміки і таргетованої метаболоміки. Крім того, для оцінки впливу зміненого під дією метформіну кишкового мікробіому на метаболізм глюкози макроорганізму людські фекальні зразки було пересаджено безмікробним мишам, а для вивчення безпосередньої взаємодії між метформіном і мікробіомом використали симулятор кишечнику in vitro.
Результати
Метформін змінює склад кишкового мікробіому
Щоб дослідити, як метформін впливає на склад кишкового мікробіому, раніше не лікованих пацієнтів з нещодавно діагностованим ЦД 2 типу рандомізували для прийому плацебо (n=18) або метформіну 1700 мг/добу (n=22) протягом 4 міс. Пацієнтам обох груп рекомендували впродовж цього періоду дотримуватись обмеженої за калоріями дієти. Через 4 міс споживання калорій зменшилось у середньому на 342 ккал/добу без значних змін між групами (р=0,90). Частину пацієнтів (n=13), які спочатку отримували плацебо, через 6 міс від початку дослідження перевели на прийом метформіну 850 або 1700 мг/добу; для валідації результатів, отриманих у рандомізованій фазі дослідження, дані цих пацієнтів аналізували після подальших 6 міс лікування.
Як і очікувалось, зважаючи на знижене споживання калорій, індекс маси тіла (ІМТ) значно зменшився в обох групах після 4 міс спостереження. Проте значне зниження концентрації глікозильованого гемоглобіну (HbA1c) і глюкози плазми натще відзначалось лише в пацієнтів, рандомізованих на прийом метформіну. Подальшого зниження ІМТ через 6 міс після переходу на метформін не відбувалось, але рівні HbA1c і глюкози плазми натще значно знизились під впливом метформіну в цій підгрупі.
Для вивчення ефектів метформіну на кишковий мікробіом було проведено секвенування всього геному за методом дробовика для 131 фекального зразка. У середньому було отримано 38 млн спарених кінцевих фрагментів для кожного зразка (від 15 до 116 млн). Таксономію і генні профілі визначали шляхом мапування високоякісних зчитувань послідовностей, що не повторюються, і за допомогою генних каталогів MEDUSA відповідно.
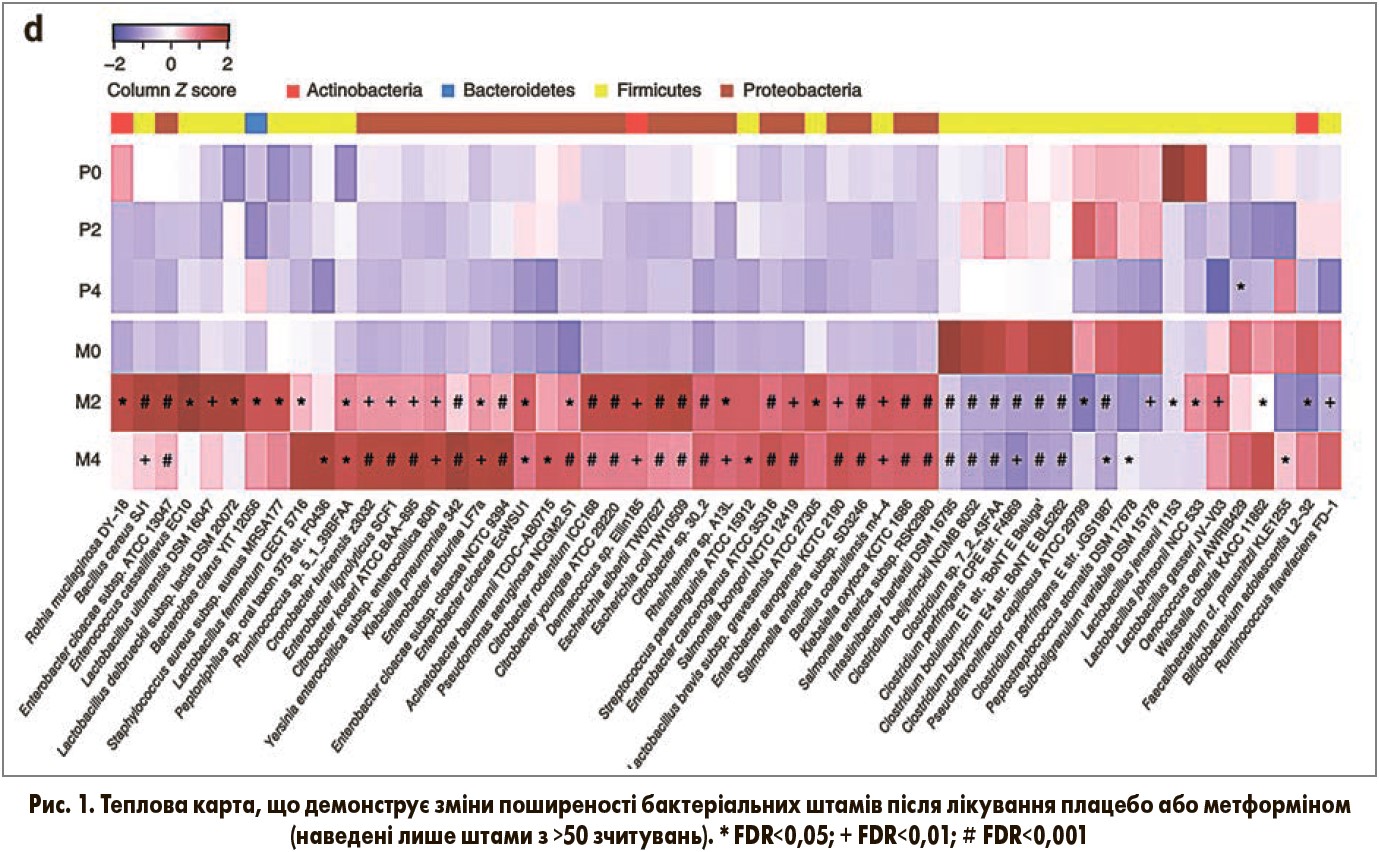 Протягом 4місячного періоду в групі плацебо змінився лише один бактеріальний штам, незважаючи на зниження ІМТ (рис. 1). Натомість після 2 і 4 міс лікування метформіном відбулись значні зміни відносної поширеності 81 і 86 бактеріальних штамів відповідно, більшість з яких належали до протеобактерій (наприклад, Escherichia coli) і Firmicutes (рис. 1). У цілому в групі метформіну спостерігали підвищення Escherichia і зниження Instestinibacter. Схожі зміни відбувались і через 6 міс у пацієнтів, яких перевели на метформін; у цій підгрупі також відзначали метформініндуковане підвищення Bifidobacterium.
Протягом 4місячного періоду в групі плацебо змінився лише один бактеріальний штам, незважаючи на зниження ІМТ (рис. 1). Натомість після 2 і 4 міс лікування метформіном відбулись значні зміни відносної поширеності 81 і 86 бактеріальних штамів відповідно, більшість з яких належали до протеобактерій (наприклад, Escherichia coli) і Firmicutes (рис. 1). У цілому в групі метформіну спостерігали підвищення Escherichia і зниження Instestinibacter. Схожі зміни відбувались і через 6 міс у пацієнтів, яких перевели на метформін; у цій підгрупі також відзначали метформініндуковане підвищення Bifidobacterium.
У попередніх експериментальних і клінічних дослідженнях був продемонстрований взаємозв’язок між метформіном і поширеністю Akkermansia muciniphila та між A. municiphila і покращенням метаболічних параметрів. Таргетований аналіз метагеномних даних показав підвищення поширеності A. muciniphila в пацієнтів, які протягом 4 міс отримували метформін. Утім цей показник не корелював зі змінами HbA1c.
Щоб встановити, як різні кишкові бактерії взаємодіють одна з одною, був проведений мережевий аналіз сумісної поширеності. Виявилось, що вже після 2 міс терапії метформіном підвищувалась кількість позитивних поєднань між родами бактерій, зокрема в межах типів Proteobacteria і Firmicutes. Також було ідентифіковано декілька зв’язків між різними типами бактерій, наприклад між Shewanella (Proteobacteria) і Blautia (Firmicutes) – родом бактерій, які продукують коротколанцюгові жирні кислоти (КЛЖК).
Для визначення впливу метформіну на ріст кишкових бактерій було проведено мапування всього геному за методом дробовика (зчитували геноми найбільш поширених кишкових мікроорганізмів людини), що дозволило порівняти співвідношення між кількістю копій ДНК біля джерела реплікації і біля точки закінчення реплікації (співвідношення «від піку до корита» – PTR) бактеріальних геномів. Після поправки на FDR (очікувана частка хибних результатів) було встановлено, що в групі метформіну показник PTR значно підвищився лише для одного виду – Bifidobacterium adolescentis; те саме спостерігалось і через 6 міс у підгрупі пацієнтів, яких перевели на метформін. Крім того, відзначалась негативна кореляція між PRT B. adolescentis і HbA1c (p<0,01). Аналіз in vitro підтвердив, що метформін безпосередньо сприяє росту B. adolescentis у чистих культурах. Також було встановлено, що в чистих культурах метформін стимулює ріст A. muciniphila, але не E. coli.
Кишковий мікробіом, змінений метформіном, покращує толерантність до глюкози
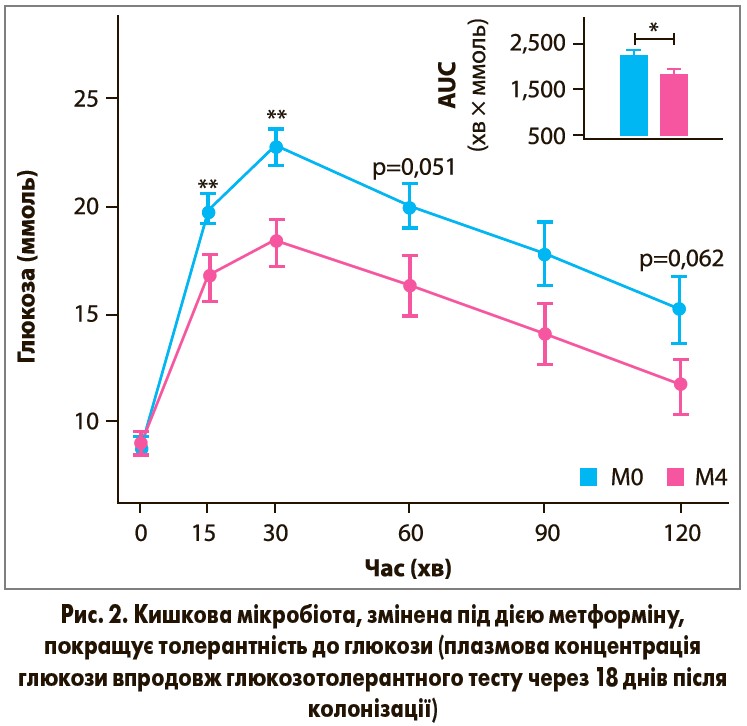 Щоб визначити, чи може змінений під дією метформіну кишковий мікробіом сприяти гіпоглікемічному ефекту препарату, фекальні зразки від трьох пацієнтів групи метформіну (до і після 4 міс лікування; мікробіоти M0 і М4) пересадили безмікробним мишам. В усіх трьох пацієнтів спостерігалась подібна відповідь на метформін стосовно зниження HbA1c через 2 і 4 міс терапії. Мишей годували їжею з високим вмістом жирів за 1 тиж до колонізації і потім ще 18 днів.
Щоб визначити, чи може змінений під дією метформіну кишковий мікробіом сприяти гіпоглікемічному ефекту препарату, фекальні зразки від трьох пацієнтів групи метформіну (до і після 4 міс лікування; мікробіоти M0 і М4) пересадили безмікробним мишам. В усіх трьох пацієнтів спостерігалась подібна відповідь на метформін стосовно зниження HbA1c через 2 і 4 міс терапії. Мишей годували їжею з високим вмістом жирів за 1 тиж до колонізації і потім ще 18 днів.
Миші, які отримали мікробіоту М0 або М4, не відрізнялись за масою тіла, вмістом жиру в тілі і рівнем інсуліну натще. Проте в реципієнтів М4 відбувалось покращення толерантності до глюкози порівняно з реципієнтами М0 (рис. 2).
Метформін викликає функціональні зсуви кишкового мікробіому
З метою подальшого вивчення функціональних змін кишкового мікробіому після лікування метформіном гени анотували в Кіотську енциклопедію генів і геномів (KEGG). У групі плацебо впродовж 4 міс значно (FDR<0,05) змінились лише два ортологи KEGG (KO). Натомість після 2 і 4 тиж лікування метформіном підвищились 626 і 472 KO та знизились 140 і 69 KO відповідно (FDR<0,05), при цьому більшість зсувів узгоджувались між двома періодами забору зразків. Аналіз головних координат відносної поширеності всіх значно змінених KO показав подібне функціонування генів у групі плацебо впродовж всього часу спостереження і в групі метформіну до початку лікування, проте після 2 і 4 тиж прийому метформіну відбувались значні зсуви. За допомогою аналізу зі збагаченням сигнальних і метаболічних шляхів було встановлено, що лікування метформіном пов’язане переважно зі збагаченням генів, які забезпечують відповіді бактерій на навколишнє середовище
(наприклад, бактеріальну систему секреції, двокомпонентну регуляторну систему і АТФзв’язувальні касетні транспортери, або АВСтранспортери), резистентність до лікарських препаратів (бактеріальний хемотаксис і катіонну антимікробну пептидну резистентність), центральний метаболізм вуглеводів (систему фосфотрансферази, метаболізм пірувату, бутирату і пропіонату), метаболізм амінокислот і біосинтез ліпополісахариду (ЛПС).
Не зовсім зрозуміло, як зміни кишкового мікробіому чинять позитивні ефекти на макроорганізм. Одним з можливих механізмів може бути підвищення продукції КЛЖК, передусім ацетату, пропіонату й бутирату, а також інших органічних кислот. Отже, щоб визначити, чи супроводжується збагачення бактеріальних генів, які відповідають за метаболізм КЛЖК, підвищенням продукції цих кислот, було використано таргетовану метаболоміку. Виявилось, що після 4 міс лікування в чоловіків спостерігалося значне підвищення фекальних концентрацій пропіонату й бутирату порівняно з групою плацебо; у жінок і в групі метформіну в цілому ця різниця була недостовірною. Також у групі метформіну після 4 міс лікування відзначалось значне підвищення фекальних концентрацій лактату і тенденція до вищих концентрацій сукцинату порівняно з групою плацебо.
Відомо, що кишкова мікробіота бере участь у регуляції метаболізму жовчних кислот (ЖК) і таким чином може впливати на метаболічні процеси макроорганізму. До того ж у низці досліджень була відзначена потенційна роль метформіну в модуляції профілю ЖК, проте цей зв’язок не надто зрозумілий. У дослідженні, що обговорюється, суттєвих змін фекальних ЖК у групі метформіну не відбувалось, проте після 4 міс терапії метформіном спостерігалось значне підвищення плазмових концентрацій ЖК (загальної, первинних, вторинних і некон’югованих) порівняно з групою плацебо. За допомогою таргетованого метагеномного аналізу було встановлено підвищення поширеності bsh – гена, який кодує гідролази жовчних солей, – після 2 міс прийому метформіну. Ці ферменти виробляються кишковою мікробіотою і каталізують декон’югацію гліцин або тауринкон’югованих ЖК. Отже, підвищення bsh може сприяти зростанню концентрації некон’югованих ЖК. Крім того, була встановлена значна негативна кореляція між концентраціями некон’югованих ЖК і HbA1c (p<0,05), що свідчить про потенційний зв’язок між модуляцією складу ЖК і терапевтичним ефектом метформіну.
Прямий вплив метформіну на кишкову мікробіоту
Для вивчення безпосередніх ефектів метформіну на кишкові бактерії фекальні зразки, отримані до початку лікування від двох донорів (13 і 49), культивували в двох окремих експериментах зі симулятором кишечнику. Ці зразки піддавали постійному впливу метформіну (10 ммоль) упродовж 1 тиж, потім профілювали мікробіоми за допомогою секвенування всього геному за методом дробовика на рівнях ДНК й РНК.
Композиційне профілювання показало, що експозиція метформіну значно змінила поширеність ДНК й РНК 24 штамів бактерій донора 13 і лише 4 штамів донора 49. Донорспецифічні ефекти метформіну включали, наприклад, підвищення поширеності РНК Bilophila wadsworthia (донор 13) і підвищення поширеності ДНК Lachnospiraceae bacterium (донор 49). A. muciniphila була єдиним таксоном, який відповів на терапію метформіном підвищенням поширеності ДНК й РНК в обох зразках, а також таксоном, поширеність якого зросла найбільше.
Одночасне функціональне профілювання метагеному й метатранскриптому показало, що експозиція метформіну значно змінює поширеність 686 і 909 KO в зразках від донорів 12 і 49 відповідно. Загалом під дією метформіну були збагачені 31 і 38 сигнальних та метаболічних шляхів (донори 13 і 49 відповідно), з яких 22 були спільними для обох зразків. Шість шляхів, які збагатились під впливом метформіну за даними метагеномного аналізу in vivo (зокрема, шляхи генів, що беруть участь у синтезі ЛПС, метаболізмі бутирату, пірувату й двокомпонентних системах), також збагачувались при експозиції метформіну в обох експериментах зі симуляцією кишечнику. Крім того, аналіз in vitro виявив збагачення метаболічних шляхів, пов’язаних з метаболізмом кофакторів і вітамінів. Ці результати свідчать, що метформін хоча й має донорспецифічні таксономічні ефекти, проте також викликає мікробні функціональні зміни, спільні для зразків від обох донорів.
Зрештою був проведений глибокий транскриптомний аналіз (з використанням культивованого in vitro зразка від донора 13) з метою визначення прямих взаємодій між метформіном і окремими штамами бактерій. Спершу були вивчені зчитування РНК, маповані в каталог генів A. muciniphila (таксон з найбільшою поширеністю в цьому фекальному зразку). Виявилось, що близько 10% (207 з 2138) протеїнкодуючих генів A. muciniphila значно змінюються під дією метформіну. Крім того, 78 з 207 метформінрегульованих генів можуть бути анотовані до KO, з них 41 ген мапується до 22 метформінзбагачених шляхів, спільних для культивованих фекальних зразків обох донорів. Шляхом ручної анотації було встановлено, що білкові продукти 108 з 207 метформінрегульованих генів потребують кофакторів або коферментів, таких як АТФ, флавінаденіндинуклеотид, флавінмононуклеотид, метали, нікотинаденіндинуклеотид і вітамін В6; більшість інших генів (69 з 99) не були охарактеризовані. Особливий інтерес викликають 81 зі 108 метформінрегульованих анотованих генів, які кодують металопротеїни або транспортери металів. Аналіз генних онтологів (GO) метформін регульованих генів A. muciniphila підтвердив, що ці продукти генів збагачені білками, які зв’язуються з іонами металів, а також з багатьма іншими кофакторами і коферментами, зокрема трансферазами, гідролазами, лігазами і білковими компонентами рибосом. Щоб визначити, чи є ці спостереження специфічними для A. muciniphila, також було проаналізовано B. wadsworthia (другий найбільш поширений таксон у цьому культивованому фекальному зразку після лікування метформіном). Згідно з детекцією гомології протеїнів лише 14 метформінрегульованих генів були ортологічними між цими двома бактеріями; проте найбільш анотовані метформінрегульовані гени B. wadsworthia також кодували металопротеїни.
Обговорення
У цьому рандомізованому плацебоконтрольованому подвійному сліпому дослідженні за участю пацієнтів з уперше діагностованим ЦД 2 типу, які дотримувалися гіпокалорійної дієти, було встановлено, що метформін, але не обмеження калорій, чинить швидкі ефекти на склад і функцію кишкової мікрофлори паралельно зі зниженням концентрацій глюкози натще і HbA1c. Пересадка фекальної мікробіоти безмікробним мишам показала, що мікробіота, змінена під дією метформіну, може покращувати метаболізм глюкози. Крім того, транскриптомний аналіз фекальних зразків, культивованих з метформіном in vitro в симуляторі кишечнику, продемонстрував, що метформін чинить прямі ефекти на мікробіоту і регулює експресію генів, які кодують металопротеїни кишкових бактерій.
Використання спарених зразків дозволило зменшити міжіндивідуальну варіабельність, властиву більшості попередніх досліджень, які вивчали вплив метформіну на кишкову мікрофлору. Також перевагою цього дослідження є те, що в ньому брали участь пацієнти з уперше діагностованим ЦД 2 типу, які раніше не приймали будьяких антигіперглікемічних препаратів, і лише невелика кількість пацієнтів у кожній групі застосовували антигіпертензивну або гіполіпідемічну терапію. Обмеження калорій жодним чином не вплинуло на отримані результати; крім того, це обмеження є відносно м’яким порівняно з попередніми дослідженнями, в яких були продемонстровані глибокі зміни кишкового мікробіому у відповідь на дієтичні втручання. Дизайн проведеного дослідження дозволив мінімізувати вплив провідних супутніх факторів, які можуть змінювати кишковий мікробіом.
Після 2 і 4 тижнів лікування метформіном спостерігались виражені зміни складу кишкових бактерій. Подібні зміни також відзначались через 6 міс у підгрупі пацієнтів, яких перевели на прийом метформіну. Зокрема, метформін стимулював ріст біфідобактерій (B. adolescentis) in vivo та in vitro. Раніше в експериментах з моделюванням метаболічного синдрому було встановлено, що поширеність B. adolescentis негативно корелює з інсулінорезистентністю. У цьому дослідженні спостерігалась негативна кореляція між B. adolescentis і концентрацією HbA1с, тобто підвищений ріст цих бактерій може сприяти протидіабетичним ефектам метформіну.
У симуляторі кишечнику єдиним таксоном, який підвищився у відповідь на метформін (на рівнях ДНК й РНК в зразках від обох донорів), була A. muciniphila, що узгоджується з попередніми дослідженнями. Ріст цього таксону в кишечнику людини, ймовірно, залежить від індивідуальних факторів, таких як вміст клітковини і поліфенолів у їжі, імунна відповідь та вік. Відсутність кореляції між поширеністю A. muciniphila і рівнем HbA1c не дозволяє зробити висновок, що ця бактерія бере участь у реалізації корисних ефектів метформіну в дослідженій когорті.
Крім складу, метформін також суттєво змінював функції кишкових мікроорганізмів, зокрема біосинтез ЛПС і КЛЖК. Підвищений біосинтез ЛПС може відображати збільшену поширеність грамнегативних бактерій, таких як Proteobacteria, проте він не супроводжувався підвищеним системним запаленням, оскільки рівень Среактивного білка не змінювався. Подібне збільшення ЛПС за відсутності підвищення системного запалення спостерігається в пацієнтів після баріатричних хірургічних втручань.
Транскриптомний аналіз показав, що більшість метформінрегульованих генів кодують металопротеїни або транспортери металів. Цікаво, що деякі метали беруть участь у патофізіології ЦД 2 типу, і упродовж багатьох років відомо, що метформін зв’язується з металами. Крім того, у нещодавньому дослідженні було встановлено, що ефекти метформіну на печінкові клітини ссавців залежать від металзв’язуючих властивостей препарату.
Пересадка безмікробним мишам фекальної мікробіоти, зміненої під впливом метформіну, забезпечила значне покращення толерантності до глюкози; це свідчить про те, що метформінадаптована мікрофлора може сприяти реалізації корисного впливу метформіну на гомеостаз глюкози. Проте слід враховувати різницю в дієтах у пацієнтів, які отримували метформін, і в тваринреципієнтів (обмеження калорій vs дієта з високим умістом жирів), що, ймовірно, посилило різницю в складі кишкової мікрофлори між донорами й реципієнтами.
Сьогодні з’являється все більше доказів, що КЛЖК і ЖК можуть брати участь у регуляції гомеостазу глюкози. Метформініндуковані зміни цих мікробнорегульованих метаболітів, які спостерігались у проведеному дослідженні, можуть частково пояснювати більш виражений гіпоглікемічний ефект метформіну в разі перорального призначення на відміну від внутрішньовенних інфузій.
Таким чином, проведене дослідження показало, що метформін взаємодіє з різними кишковими бактеріями (можливо, шляхом регуляції гомеостазу металів). Для ідентифікації інших мікробних метаболітів або білків і визначення того, як вони покращують метаболізм макроорганізму, необхідні додаткові дослідження, що поєднують нетаргетовану метаболоміку й метапротеоміку.
Список літератури знаходиться в редакції.
Стаття друкується в скороченні.
Wu H. et al. Metformin alters the gut microbiome of individuals with treatmentnaive type 2 diabetes, contributing to the therapeutic effects of the drug. Nat Med. 2017 Jul; 23(7): 850858.
Переклав з англ. Олексій Терещенко
Медична газета «Здоров’я України 21 сторіччя» № 15-16 (412-413), серпень 2017 р.
СТАТТІ ЗА ТЕМОЮ Терапія та сімейна медицина
Як відомо, кальцій бере участь у низці життєво важливих функцій. Хоча більшість досліджень добавок кальцію фокусувалися переважно на стані кісткової тканини та профілактиці остеопорозу, сприятливий вплив цього мінералу є значно ширшим і включає протидію артеріальній гіпертензії (передусім у осіб молодого віку, вагітних та потомства матерів, які приймали достатню кількість кальцію під час вагітності), профілактику колоректальних аденом, зниження вмісту холестерину тощо (Cormick G., Belizan J.M., 2019)....
Після десятиліть, а часом і запеклих суперечок про переваги та недоліки застосування глюкокортикоїдів (ГК) досягнута певна конвергенція. Сучасні рекомендації лікування таких захворювань, як ревматоїдний артрит (РА), ревматична поліміалгія (РПМ) та васкуліт великих судин відображають поточний стан консенсусу терапії ГК. Однак залишаються відкритими питання щодо можливості тривалого лікування дуже низькими дозами ГК у пацієнтів із РА, а також успішності пошуку інноваційних ГК (лігандів ГК-рецепторів) із покращеним співвідношенням користь/ризик....
Серед препаратів, які мають велику доказову базу щодо лікування пацієнтів із захворюваннями дихальних шляхів з алергічним компонентом, особливий інтерес становлять антагоністи лейкотрієнових рецепторів (АЛТР). Ці препарати мають хорошу переносимість у дорослих та дітей, а також, на відміну від інгаляційних кортикостероїдів (ІКС), характеризуються високим комплаєнсом, тому посідають чільне місце в лікуванні пацієнтів із респіраторною патологією. У лютому відбувся міждисциплінарний конгрес із міжнародною участю «Життя без алергії International» за участю провідних вітчизняних міжнародних експертів. Слово мав президент Асоціації алергологів України, професор кафедри фтизіатрії та пульмонології Національного університету охорони здоров’я України ім. П.Л. Шупика (м. Київ), доктор медичних наук Сергій Вікторович Зайков із доповіддю «Місце АЛТР у лікуванні пацієнтів із респіраторною патологією». ...
Розбір клінічного випадку...